Todos los músculos esqueléticos están formados por numerosas fibras cuyo diámetro varía de 10 y 80 m.
Cada fibra muscular contiene va rios cientos a varios miles de miofibrillas y cada miofibrilla está formada por aproximadamente 1.500 filamentos de miosina y 3.000 filamentos de actina.
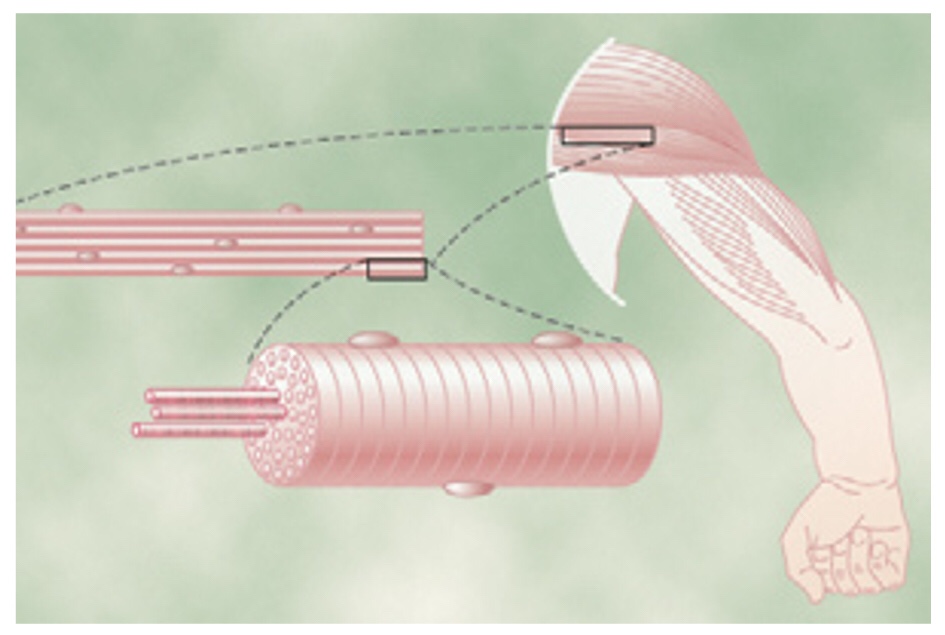
Los filamentos de miosina y de actina se interdigitan parcialmente y de esta manera hacen que las miofibrillas tengan bandas claras y oscuras alternas.
Las bandas claras contienen sólo filamentos de actina y se denominan bandas I porque son isótropas a la luz polarizada. Las bandas oscuras contienen filamentos de miosina, y se denominan bandas A porque son anisótropas a la luz polarizada.

Las pequeñas proyecciones que se originan en los lados de los filamentos de miosina se denominan puentes cruzados. La interacción entre estos puentes cruzados y los filamentos de actina produce la contracción.
Los extremos de los filamentos de actina están unidos al denominado disco Z. El disco Z, que en sí mismo está formado por proteínas filamentosas, pasa desde unas miofibrillas a otras, uniéndolas entre sí a lo largo de toda la longitud de la fibra muscular.
Por tanto, toda la fibra muscular tiene bandas claras y oscuras, al igual que las miofibrillas individuales. Estas bandas dan al músculo esquelético y cardíaco su aspecto estriado.
La porción de la miofibrilla (o de la fibra muscular entera) que está entre dos discos Z sucesivos se denomina sarcómero.
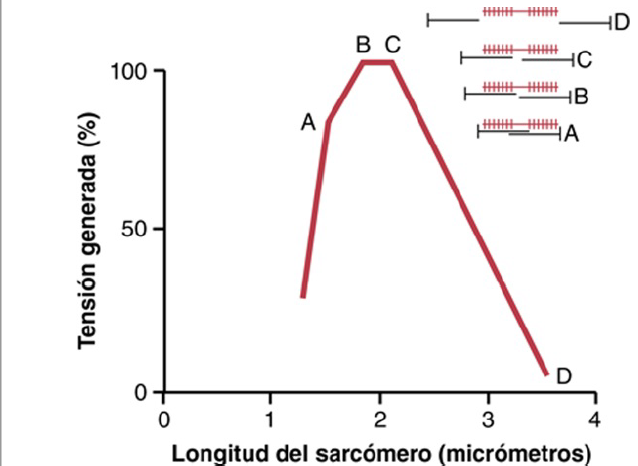
Cuando el sarcómero tiene una longitud, de 2m. los filamentos de actina se superponen completamente con los filamentos de miosina y las puntas de los filamentos de actina están comenzando ya a superponerse entre sí.
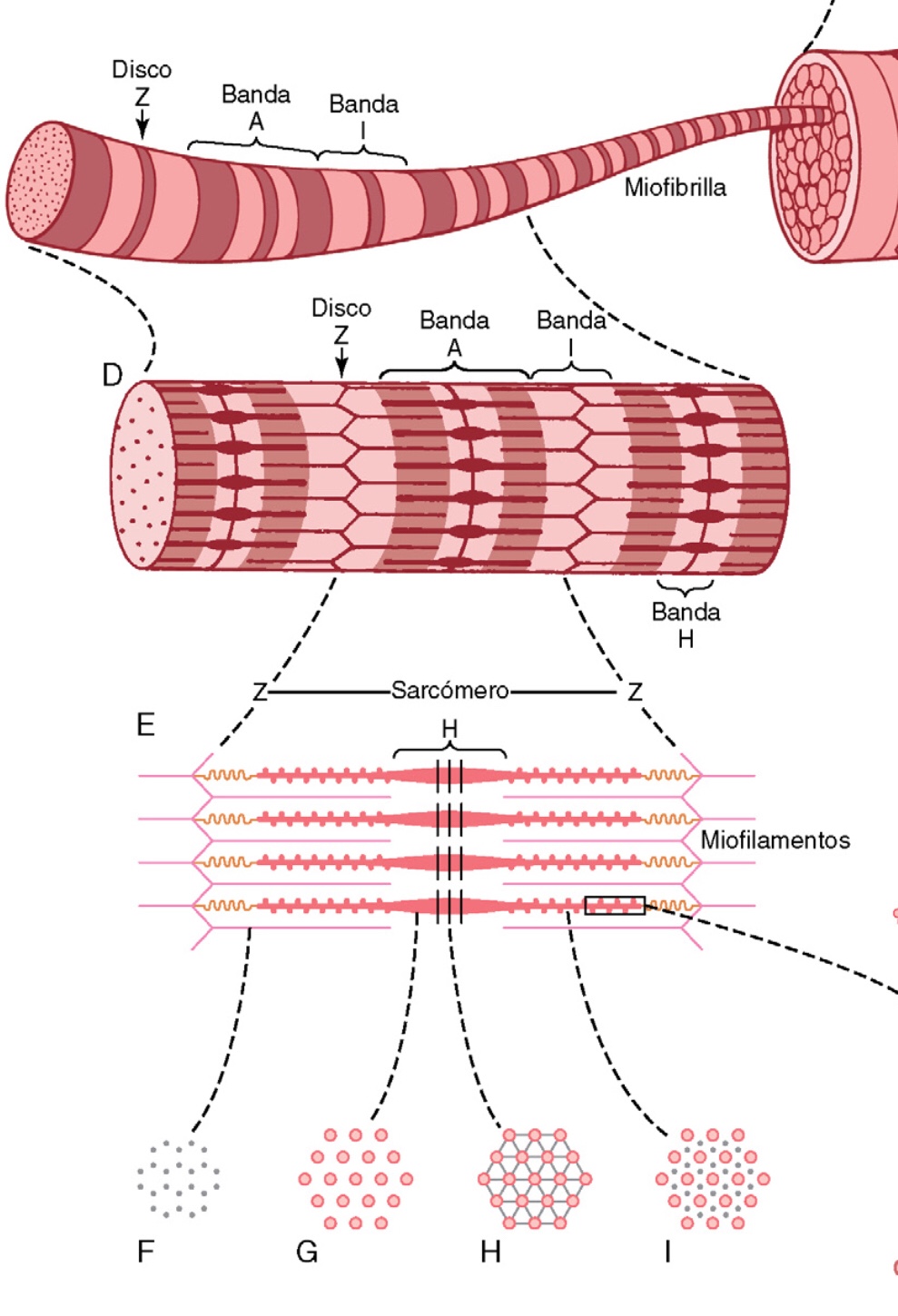
Las moléculas filamentosas de titina mantienen en su lugar los filamentos de miosina y actina.
Las bandas claras contienen solo filamentos de actina y se denominan bandas I porque son
isótropas a la luz polarizada. Las bandas oscuras contienen filamentos de miosina, así como los extremos de los filamentos de actina en el punto en el que se superponen con la miosina, y se denominan bandas A porque son anisótropas a la luz polarizada.
Los espacios entre las miofibrillas están llenos de un líquido intracelular denominado sarcoplasma, que
contiene grandes cantidades de potasio, magnesio y fosfato, además de múltiples enzimas proteicas.
También hay muchas mitocondrias que proporcionan energía en forma ATP.

En el sarcoplasma que rodea a las miofibrillas de todas las fibras musculares también hay un extenso
retículo sarcoplásmico, tiene una organización especial que es muy importante para regular el almacenamiento, la liberación y la recaptación de calcio.

En el estado relajado, los extremos de los filamentos de actina que se extienden entre dos discos Z comienzan a superponerse entre sí.
Por el contrario, en el estado contraído estos filamentos de han sido traccionados hacia dentro entre los filamentos de miosina, de modo que sus extremos se superponen entre sí en su máxima extensión.
Así, la contracción muscular se produce por un mecanismo de deslizamiento de los filamentos.
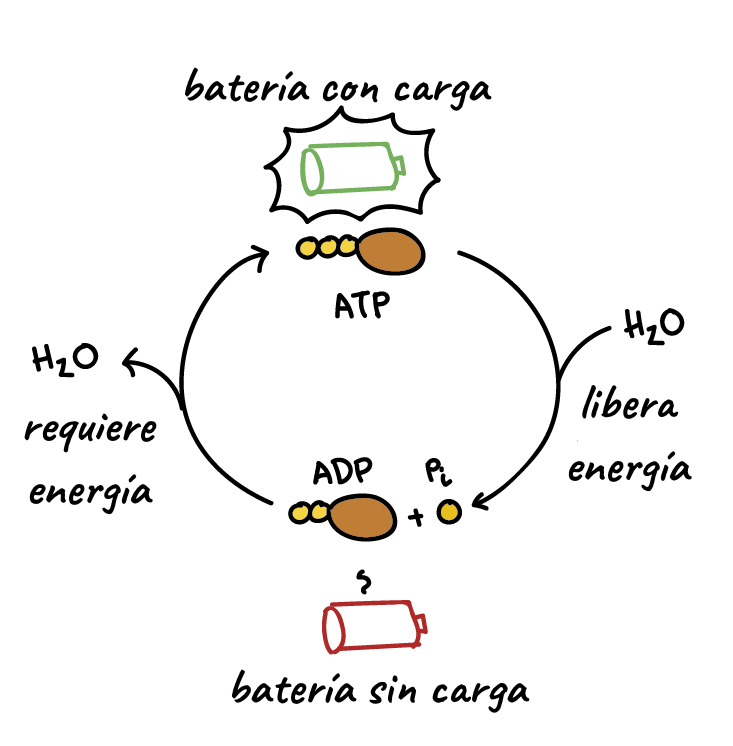
Sin embargo, es necesaria energía para que se realice el proceso contráctil. Esta energía procede de los enlaces de alta energía de la molécula de ATP, que es degradada
a ADP.
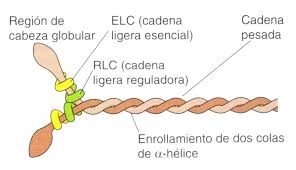
La molécula de miosina está formada por seis cadenas polipeptídicas, dos cadenas pesadas y cuatro cadenas
ligeras. Las dos cadenas pesadas se enrollan entre sí en espiral para formar una hélice doble, que se denomina cola de la molécula de miosina.
Un extremo de cada una de estas cadenas se pliega para formar una estructura
polipeptídica globular denominada cabeza de la miosina. Así, hay dos cabezas libres en un extremo de la
molécula de miosina de doble hélice.
Cada una de las hebras de la doble hélice de F-actina está formada por moléculas de G-actina. A cada una
de estas moléculas se le une una de ADP.
El filamento de actina también contiene otra proteína, la tropomiosina. Estas moléculas están enrolladas en espiral alrededor de los lados de la hélice de F-actina.

Los brazos y las cabezas que protruyen se
denominan en conjunto puentes cruzados. Cada puente cruzado es flexible en dos puntos denominados bisagras, una en el punto en el que el brazo sale del cuerpo del filamento de miosina y la otra en el punto en el que la cabeza se une al brazo.

En estado de reposo las moléculas de tropomiosina
recubren los puntos activos de actina, de modo que no se puede producir atracción entre los filamentos de actina y de miosina para producir la contracción.
Troponina y su función en la contracción muscular
A lo largo de los lados de las moléculas de tropomiosina hay otras moléculas proteicas denominadas troponina. Estas moléculas proteicas son en realidad complejos de
tres subunidades proteicas unidas entre sí.
- La troponina I tiene una gran afinidad por la actina.
- La troponina T por la tropomiosina
- La troponina C por los iones calcio.
Este complejo une la tropomiosina a la actina y la afinidad de la troponina por los iones calcio inicia el proceso de la contracción.

A medida que el sarcómero se acorta y que el filamento de actina comienza a superponerse al filamento de
miosina, la tensión aumenta progresivamente hasta que la longitud del sarcómero disminuye a 2,2 μm.
Con un acortamiento adicional los extremos de los dos filamentos de actina comienzan a superponerse entre sí además de superponerse a los filamentos de miosina, la longitud del sarcómero disminuye hasta 1,65 μm.
Cuando se produce la contracción, los extremos de los
filamentos de miosina están corrugados y la fuerza de la contracción se aproxima a cero, aunque el sarcómero ya se ha contraído hasta su mínima longitud.
Tres fuentes de energía para la contracción muscular
La primera fuente de energía que se utiliza para reconstituir el ATP es la sustancia fosfocreatina, que
contiene un enlace fosfato de alta energía similar a los enlaces del ATP.

La energía combinada del ATP y de la fosfocreatina en el músculo es capaz de producir una contracción muscular máxima durante solo 5 a 8 s.
La segunda fuente importante de energía es la «glucólisis» del glucógeno. La escisión del glucógeno en ácido pirúvico y láctico libera energía. Después se puede utilizar el ATP para aportar energía a la contracción o para reconstituir los almacenes de fosfocreatina.
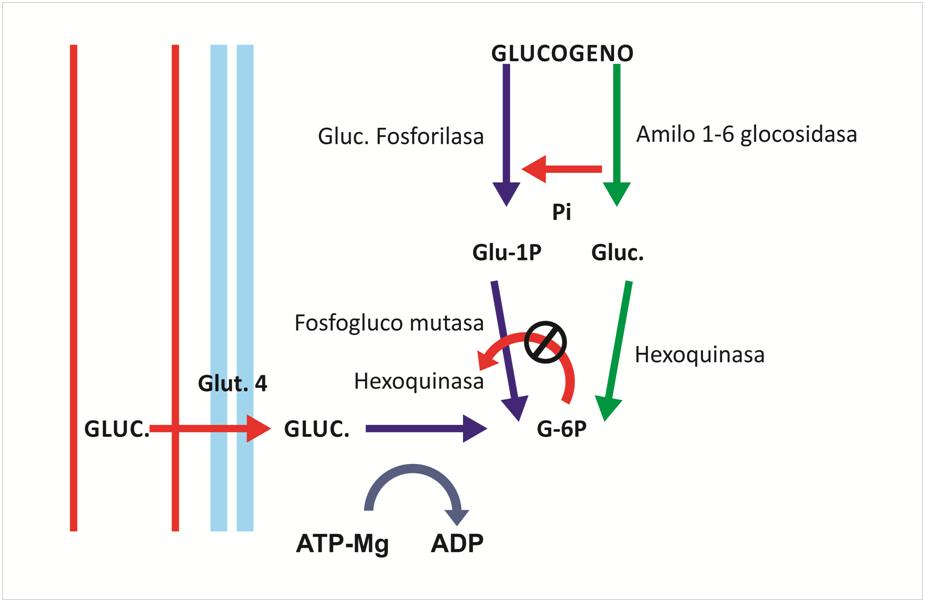
La importancia de este mecanismo es:
- Pueden producir incluso en ausencia de oxígeno, de modo que se puede mantener la contracción muscular durante muchos segundos (más de 1 min).
- La velocidad de formación de ATP por el proceso es más rápida.
- Sin embargo, se acumulan tantos productos finales que pierde su capacidad de mantener una contracción máxima después de 1 min.
La tercera y última fuente de energía es el metabolismo oxidativo, lo que supone combinar oxígeno con
los productos finales de la glucólisis y con otros diversos nutrientes celulares para liberar ATP.
Más del 95% de toda la energía que utilizan los músculos para la contracción sostenida a largo plazo procede del metabolismo oxidativo.
- Actividad muscular máxima a muy largo plazo (muchas horas) lla energía procede con mucho de las grasas,
- Durante períodos de 2 a 4 h hasta la mitad de la energía puede proceder de los hidratos de carbono almacenados.

Sistemas de palanca del cuerpo
En resumen, el análisis de los sistemas de palanca del cuerpo depende del conocimiento de:
- 1) El punto de la inserción muscular
- 2) Su distancia desde el fulcro de la palanca
- 3) La longitud del brazo de la palanca
- 4) La posición de la palanca.

En el cuerpo son necesarios muchos tipos de movimiento,
algunos de los cuales precisan una intensidad grande, y otros precisan grandes distancias de movimiento.
Por este motivo hay muchos tipos diferentes de músculo.

El estudio de los diferentes tipos de músculos, de los
sistemas de palanca y de sus movimientos se denomina cinesiología y es un componente científico
importante de la fisioanatomía humana.
Hipertrofia y atrofia muscular
El aumento de la masa total de un músculo se denomina hipertrofia muscular. Cuando la masa total
disminuye, el proceso recibe el nombre de atrofia muscular.
Toda la hipertrofia muscular se debe a un aumento del número de filamentos de actina y miosina en cada fibra muscular, dando lugar a aumento de tamaño de las fibras musculares individuales; esta situación se denomina hipertrofia de las fibras.
Cuando un músculo no se utiliza durante muchas semanas, la velocidad de degradación de las proteínas
contráctiles es mucho más rápida que la velocidad de sustitución. Por tanto, se produce atrofia muscular.

Otro tipo de hipertrofia se produce cuando los músculos son distendidos hasta una longitud mayor de lo normal. Esta distensión hace que se añadan nuevos sarcómeros en los extremos de las fibras musculares, donde se unen a los tendones.
Por el contrario, cuando un músculo permanece acortado a una longitud menor que su longitud normal de manera continua, los sarcómeros de los extremos de las fibras musculares pueden llegar realmente a desaparecer.
Hiperplasia de las fibras musculares
En situaciones de generación extrema de fuerza muscular se observa un aumento real del número de fibras musculares (además de la hipertrofia de las fibras). Este aumento del número de fibras se denomina hiperplasia de las fibras.
En la fase final de la atrofia por denervación, la mayor parte de las fibras musculares es destruida y
sustituida por tejido fibroso y adiposo. Las fibras que permanecen poseen propiedades contráctiles
escasas o nulas.

El tejido fibroso que sustituye a las fibras musculares durante la atrofia por denervación también
tiende a seguir acortándose durante muchos meses, lo que se denomina contractura.
Recuperación de la contracción muscular en la poliomielitis
Cuando se destruyen algunas fibras nerviosas que inervan un músculo, pero no todas, como ocurre con
frecuencia en la poliomielitis.

Este proceso da lugar a unidades motoras de gran tamaño denominadas macrounidades motoras, que pueden contener hasta cinco veces el número normal de fibras musculares para cada neurona que procede de la médula espinal.
Rigidez cadavérica
Varias horas después de la muerte, todos los músculos del cuerpo entran en un estado de contractura denominado «rigidez cadavérica»; es decir, los músculos se contraen y se hacen rígidos, incluso sin potenciales de acción.
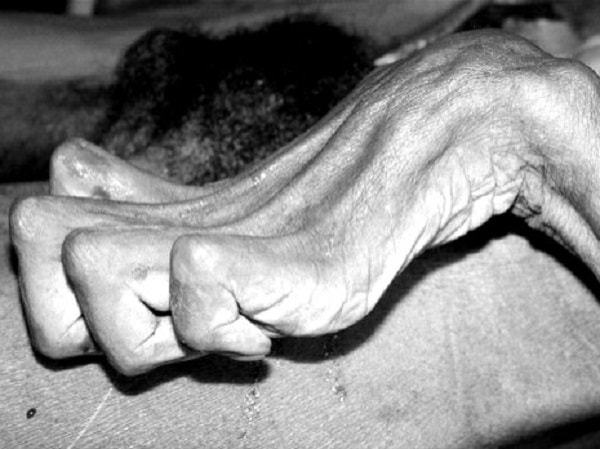
Esta rigidez se debe a la pérdida de todo el ATP, necesario para producir la separación de los puentes cruzados durante el proceso de relajación.
Distrofia muscular
Comprenden varios trastornos hereditarios susceptibles de causar debilidad y degeneración progresiva de las fibras musculares, que son sustituidas por tejido graso y colágeno.
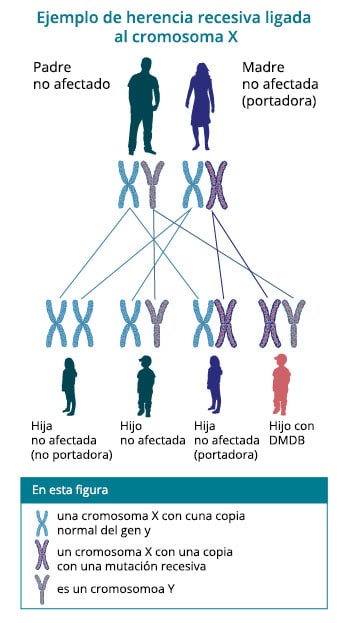
Una de las formas más comunes de distrofia muscular es la distrofia muscular de Duchenne (DMD).

Esta enfermedad afecta solo a los hombres, debido a que se transmite como un rasgo recesivo ligado al
cromosoma X y es provocada por la mutación de un gen que codifica para la distrofina.
La falta de distrofina o formas mutadas de la proteína provocan desestabilización de la membrana de las células de los músculos, activación de múltiples procesos fisiopatológicos y dificultad en la reparación de las membranas deterioradas después de un daño.
- Debilidad muscular que comienza en la primera infancia y avanza rápidamente
- Los pacientes suelen tener que desplazarse en silla de ruedas desde los 12 años de edad
- Mueren a consecuencia de un fallo respiratorio antes de cumplir los 30.

Una forma más leve de esta enfermedad, denominada distrofia muscular de Becker (DMB), tiene un inicio más tardío e índices más elevados de supervivencia.
Se calcula que la DMD y la DMB afectan a 1 de cada 5.600 a 7.700 hombres de entre 5 y 24 años de edad.

En la actualidad no existe ningún tratamiento eficaz contra la DMD o la DMB, aunque se abre la posibilidad de recurrir a la terapia génica en el futuro.
